Servicio de Cardiología. Hospital Ramón y Cajal. Madrid
1. Las enfermedades cardiovasculares
La primera causa de muerte en el mundo occidental son las enfermedades cardiovasculares y se cree que irán en ascenso en todo el mundo, al aumentar el envejecimiento general de la población y al implantarse los hábitos de la vida moderna en los países emergentes con menores recursos. Dentro de ellas, las debidas a cardiopatía coronaria (en adelante CC), las vasculo-cerebrales y las vasculares periféricas son las responsables del 60% del total de mortalidad. En este artículo nos centraremos sobre todo en las primeras.
El fenómeno fisiopatológico común y subyacente a estas enfermedades es el proceso denominado «aterosclerosis». Este proceso comienza en la juventud, con el depósito graso en la pared arterial, llamado placa de ateroma que, en principio, es asintomático, hasta que se manifiesta después en edades avanzadas de la vida, con el fenómeno de «aterotrombosis», que si ocurre en las arterias coronarias origina un infarto de miocardio, o bien un ictus si aparece en las arterias cerebrales o carotídeas (Figura 1).
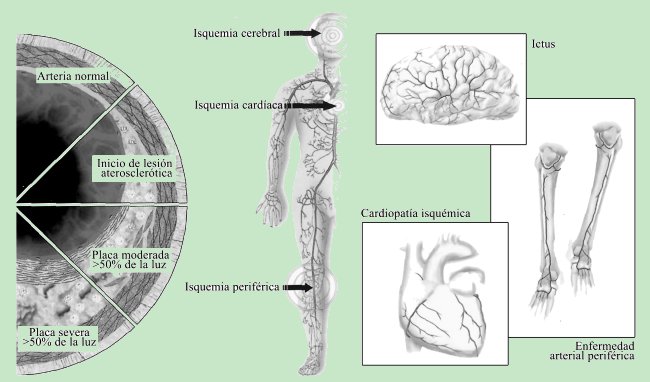 |
| Figura 1. Evoluciones de la placa y manifestaciones clínicas de la aterosclerosis. La aterosclerosis puede conducir al desarrollo de complicaciones graves. El infarto de miocardio, el ictus y la enfermedad vascular periférica son las principales consecuencias de esta afección. |
La aterosclerosis se produce por la interacción de factores ambientales y genéticos adversos. Se cree que tienen mayor importancia los factores ambientales como el tabaquismo y los malos hábitos de vida, como la dieta desequilibrada, el consumo de alcohol, la falta de ejercicio físico etc., como indican los estudios de diferentes poblaciones y de emigrantes en diversos países. Además, se han identificado distintos factores de riesgo clásicos como la hipercolesterolemia, la hipertensión, la resistencia insulínica etc. presentes en la etiopatogenia de la arteriosclerosis, aunque estos factores no explican la variabilidad encontrada en la incidencia de CC en diversas poblaciones.
Por lo que se refiere a los factores genéticos, al igual que en otras enfermedades comunes de la vida adulta, son prevalentes en la población pero con baja penetrancia, por lo que el componente genético en solitario explica sólo un pequeño porcentaje del riesgo de CC. Así, aunque por ahora se han identificado más de 50 genes con posible influencia sobre las CC, fundamentalmente los del metabolismo lipoproteico, como los de las apolipoproteínas y los del sistema renina-angiotensina (sustancias con actividad vasoconstrictora), es muy rara la susceptibilidad originada como resultado de una única mutación génica. Las excepciones serían casos como la hipercolesterolemia debida a una mutación del receptor de la lipoproteína de baja densidad (LDLc-colesterol malo) o a la mutación de arginina-glutamina (Arg-3,500-Glu) de la apoliproteína B (lipoproteína del LDLc), que están presentes en cerca de 1 de cada 500 y de 1 cada 1000 sujetos respectivamente, causando CC por sí solas.
La aterosclerosis es, por tanto, una enfermedad multifactorial que se produce por la interacción de diversos factores ambientales, también llamados factores de riesgo clásicos, y los factores de riesgo genotípicos. Entre los dos grupos de factores, se mantendría una estrecha relación e interacción durante toda la vida, pero sus efectos negativos se manifiestan sobre todo en la edad adulta post-reproductiva de la humanidad.
Sin embargo, en la actualidad persisten muchas dudas sobre las causas y los efectos de las cardiopatías coronarias. En este trabajo se revisa la situación existente con respecto a algunos de estos aspectos y, sobre todo, se trata de explicar, utilizando un enfoque darwiniano-evolucionista, algunos de los enigmas existentes con respecto a la baja incidencia de estas enfermedades en determinadas regiones o poblaciones.
2. Teorías sobre el origen de la aterosclerosis
En 1956, Sinclair promovió la idea de que la alta incidencia de CC era debida a la gran ingesta de colesterol y grasas saturadas, lo que producía un aumento del colesterol plasmático total. La presencia de células espumosas (macrófagos) cargadas de colesterol en las placas de ateroma, dio la base a la hipótesis lipídica o de dieta-lípidos-corazón. Los estudios epidemiológicos de los Siete Países (A. Keys, 1986) y de las migraciones de diferentes grupos étnicos también apoyaban esta idea.
Esta hipótesis se completó con la de respuesta a la lesión de R. Ross (1990), donde se implicaban factores inflamatorios, como los monocitos-macrófagos y ciertos subconjuntos de las células T (CD4+ y CD8+), en la respuesta endotelial arterial. Sin embargo, estas hipótesis sólo explican el 50-60% de la incidencia de la cardiopatía coronaria (CC).
En 1989, A. Steinberg y J. Witzum propusieron la hipótesis antioxidante, que postulaba que era imprescindible que las lipoproteínas de baja densidad (LDLc-colesterol malo) estuviesen modificadas, básicamente oxidadas (LDLc-ox), para ser aterogénicas.
Posteriormente, se ha desarrollado por G. Wick (2001), la llamada hipótesis inmunológica de la aterosclerosis, centrada en los estadios iniciales de la enfermedad y que parece completar al resto de teorías mencionadas. Esta hipótesis propugna que una chaperona-proteína que «ayuda» a funcionar a otras proteínas, la proteína del choque térmico 60 (HSP-60), implicada en la defensa inmunológica, puede promover la aterosclerosis, en respuesta al estrés endotelial inducido por los factores de riesgo clásicos.
Sin embargo, quedan importantes dudas por resolver para entender todo el proceso aterosclerótico y en concreto en relación con su incidencia en distintas poblaciones humanas. Y tal como se dice en la introducción, se va a tratar de explicar, sobre todo, este último aspecto dentro del marco de lo que se llama Medicina Evolucionista.
3. La medicina evolucionista
La medicina, esta basada en la Biología, y dentro de ésta, la Evolución es su teoría explicativa principal. Y sin embargo, aunque uno de los padres de la moderna teoría de la evolución, Theodosius Dobzhanky (1970), afirmó que: «Nada tiene sentido en biología, salvo a la luz de la evolución», la medicina ha permanecido claramente separada de la evolución, durante más de cien años. Esta ignorancia, dada la naturaleza científica de la medicina, sorprende. No parece que se deba a las críticas creacionistas a la selección natural. Tal vez esta desatención, se deba a la idea de que en alguna forma el progreso médico neutraliza la influencia de la selección natural. Pero, precisamente, la medicina evolucionista trata de comprender esta influencia. En este sentido, la medicina evolucionista se define como: «el estudio de las consecuencias rápidas y dinámicas de la selección natural sobre las adaptaciones del linaje humano y sus patologías, así como la reconstrucción de su historia evolutiva y sus consecuencias para la salud y la enfermedad». Con el comienzo del nuevo milenio, la medicina evolucionista se está convirtiendo en una nueva disciplina que aplica un enfoque darwiniano a las enfermedades humanas y a sus causas y sus remedios. De hecho, dentro de este campo existen una serie de principios básicos que se van a exponer primero en términos generales, y a continuación en relación con las enfermedades cardiacas.
Así, la medicina evolucionista reconoce, en primer lugar, que algunos problemas de salud de la humanidad pueden ser debidos a problemas en el diseño de nuestro cuerpo resultantes de los procesos evolutivos a los que nos hemos visto sometidos. Por ejemplo, la existencia de una zona ciega de la retina, es característica del particular desarrollo del ojo de los mamíferos y el hipo, es un vestigio de nuestro pasado acuático. En este contexto es posible considerar que parte de los variantes -alelos- que posee el genoma de la humanidad son el resultado de la adaptación al ambiente ancestral de la humanidad -por ejemplo a dietas «naturales» del Paleolítico y/o del Neolítico- y que, ahora, con el paso del tiempo, nuestro ambiente físico y cultural es diferente -la dieta, concretamente- con lo que alelos que en otro tiempo fueron favorables ahora pueden ser desfavorables –al menos en ciertos ambientes- por lo que está aumentando la frecuencia de ciertas enfermedades como la diabetes, la obesidad, la hipertensión y también las enfermedades cardiacas.
Un ejemplo típico de este tipo de cambios, es lo que ha pasado en la humanidad en tiempos recientes con la intolerancia a la lactosa en edades adultas. Y es que el hombre, como los mamíferos en general, es capaz de degradar la lactosa de la leche en el periodo de lactancia (por la acción de un enzima llamada lactasa) pero, y en esto es excepcional, también en las etapas adultas. Esta capacidad nueva de la especie humana se adquirió en principio en las sociedades donde comenzó la ganadería de producción de leche (es decir en el Neolítico) merced a que se seleccionaron los genotipos que producen lactasa después del periodo natural de lactancia. Pero mientras sucedió esto debió de haber muchos casos de muertes y enfermedades por intolerancia a la lactosa, y de hecho todavía hay casos y sociedades donde es frecuente este problema.
En segundo lugar, se ha comprobado que puede haber situaciones en que un determinado variante-alelo de un cierto gen puede ser causante de una enfermedad en homocigosis, pero que en heterocigosis confiere a los individuos que lo portan superior valor adaptativo -a esto se le llama ventaja de los heterocigotos. Esto ocurre, por ejemplo, en la anemia falciforme, en la fibrosis quística o en la fenilcetonuria. En el caso de la anemia falciforme lo que sucede es que es frecuente en poblaciones -como en África- donde la malaria es endémica, y eso es debido a que en dichas regiones el genotipo heterocigótico para el alelo que produce la anemia es ventajoso (en la sangre de los individuos portadores de la mutación no se completa bien el ciclo de vida del protozoo que causa la malaria) comparado con los individuos homocigóticos normales para el gen de la anemia (en ellos se completa bien el ciclo de vida del parásito) y, por supuesto, en comparación con los homocigóticos para el alelo mutante que tienen una grave anemia.
Asimismo, en tercer lugar se está empezando a comprender que hay características de la humanidad que son el resultado de la selección natural, por las ventajas que reúnen hasta la reproducción, pero que dichas características pueden ser desfavorables y dar serios problemas de salud en épocas post-reproductivas más tardías de la vida. Esto es lo que se ha venido a llamar por Haldane en 1950, pleitropía antagónica, que se refiere: 1º) a cuando un gen afecta a dos o más rasgos, uno ventajosamente evolutivo y otro deletéreo. 2º) cuando un rasgo establecido por selección natural, es ventajoso hasta la edad de la reproducción, pero puede llegar a ser adverso en la vida tardía. Esto es lo que por ejemplo ocurre con el gen Snail 1, que controla la mineralización ósea en la juventud y por tanto protege de las fracturas. Hay un alelo en este gen, que es regulador positivo de la beta-catenina, que actúa sobre el receptor de la vitamina D (RVD) y que aumenta la mineralización, por lo que al mismo tiempo que hace que sus poseedores se reproduzcan más que los que carecen de él, a cambio produce una mayor calcificación arterial, aterosclerosis y neoplasias de colon en edades avanzadas de la vida. Por tanto, este variante génico a pesar de significar mayor riesgo en la salud de los individuos mayores, estará presente en el genoma de sus descendientes por su efecto positivo hasta la edad reproductiva. También se cree que algo similar puede ocurrir en la hemocromatosis, la gota, el Alzheimer, la hipertrofia prostática, etc.
4. Diferencias en CC entre poblaciones o regiones
La cardiopatía coronaria no está homogéneamente distribuida en todas las regiones y poblaciones donde habita la humanidad en un régimen de «vida moderna». Así, en Europa hay un gradiente Norte/Sur para las CC con las mayores incidencias en Finlandia y en el Reino Unido y las más bajas en la Península Ibérica y Francia. Incluso la incidencia, puede variar hasta en cuatro veces entre el Norte y el Sur de Europa; por ejemplo, 835 casos de CC por 100.000 habitantes en Karelia del Norte (Finlandia), o 777 en Glasgow (Escocia), frente a los 233 en Toulouse ( Sur de Francia ), los 210 en Gerona (España), 258 en Sicilia (Italia) y los 246 en Attica (Grecia).
Esta diferencia no tiene una clara explicación científica, ya que los factores de riesgo clásicos no sólo son parecidos en las poblaciones mediterráneas actuales a los de las otras poblaciones occidentales, sino que ya estaban presentes hace más de 50 años. Esta menor incidencia de las enfermedades cardiacas en los países mediterráneos se achacó inicialmente al alto consumo en ellos de aceite de oliva, que es muy rico en ácido oleico y que ofrece una alta resistencia a la oxidación, lo que puede favorecer la estabilidad de las LDLc. Sin embargo, países mediterráneos como Francia consumen poco aceite de oliva y bastante grasa saturada, manteniendo una baja incidencia de CC, hecho conocido como «paradoja francesa» y atribuido parcialmente al consumo de vino tinto -rico en un antioxidante como el resveratrol. Y, por otro lado, esta paradoja francesa-mediterránea se ha tratado de explicar por la dieta mediterránea en general, caracterizada por el consumo de verduras frescas escasamente procesadas y frutas con alto contenido de vitaminas y micronutrientes antioxidantes. Recientemente se ha tratado de explicar esta paradoja en relación con el tamaño de los telómeros (M. Blasco, 2007). Así, entendiendo que la aterosclerosis y su estadio final, la rotura de placa arterial (aterotrombosis) es un proceso en el que está implicado el envejecimiento y la replicación celular, y que esto, a su vez, depende del tamaño de los telómeros celulares, se ha planteado la hipótesis de que sería el tamaño de estas zonas terminales de los cromosomas, lo que podría explicar esta paradoja. De hecho, se ha comprobado que la población francesa para similares rangos de edad tiene un tamaño de telómeros celulares más largos que el resto de poblaciones europeas. Pero esto mismo no se ha encontrado en el resto de las poblaciones mediterráneas.
Por tanto, si los factores de riesgo clásicos conocidos, como la hipertensión, la diabetes, el tabaquismo, la hipercolesterolemia, etc, u otros descubiertos más recientemente -como el tamaño de los telómeros- no han sido capaces de explicar la distinta incidencia de CC entre los países del Norte y Sur de Europa, esto sugeriría que podrían existir otros factores de protección desconocidos, ya sea ambientales o genéticos, y/o que la interacción entre ellos podría explicarla. Además hay poblaciones singulares, con baja incidencia de enfermedades cardiovasculares, como, por ejemplo, las comunidades Amish de Estados Unidos (EE UU) -véase 5.2- en las que tampoco los tradicionales factores de riesgo clásicos explicarían totalmente esa baja incidencia.
5. Un enfoque evolucionista de las enfermedades cardíacas
El patrón actual paradójico, de incidencia de enfermedades cardiovasculares en poblaciones como las Europeas o las de los Amish de EE UU, podría deberse a la historia evolutiva de estas poblaciones, por lo que, aprovechando la emergencia de la medicina evolucionista y darwiniana, intentaremos dar una explicación dentro de este marco conceptual a estas paradojas y a estas observaciones.
5.1. Las enfermedades cardiacas y los cambios en la dieta de las poblaciones europeas
Se cree que gran parte de la pandemia actual de las enfermedades cardiovasculares se debe a la transición desde dietas y modos de vida más naturales de nuestros antecesores, hacia una dieta opulenta y un modo de vida sedentario y desequilibrado, derivados ambos del desarrollo económico de las sociedades occidentales actuales. Este enfoque ha dado lugar a los conceptos del genotipo ahorrador o al paradigma del mono obeso y su correlato fisiopatológico denominado «síndrome metabólico», caracterizado por: obesidad, hipertensión, resistencia insulínica, hiperlipemia mixta, sedentarismo, estrés oxidativo y enfermedad cardiovascular.
La etapa preagrícola (Paleolítico) resultó fundamental para la configuración de nuestro genoma actual. Nuestra especie desde entonces posee una dotación genética con una gran eficiencia para la acumulación de grasa en épocas de bonanza alimentaria, debido a que se modeló con escasez crónica de alimentos y con un enorme esfuerzo físico invertido en obtenerlos. Para este genotipo, J Neel en Minnessota (EE UU) en 1962, acuñó la expresión «genotipo ahorrador».
La aparición y desarrollo de la agricultura (Neolítico) transformó los patrones alimentarios y la presión selectiva se relajó hasta llegar a nuestros días. Por tanto, en las condiciones actuales de abundancia, con gran ingesta calórica total y poco gasto energético para obtenerla, los genotipos ahorradores presentes en el acervo genético humano actual desde el Paleolítico dejan de hallarse en armonía con el entorno y, por ello, se produce un aumento de «enfermedades» entre las que figuran las cardiacas.
En relación con las enfermedades cardiacas, como ejemplo específico y particular de cambios en genes relacionados con la dieta estarían los genes que interaccionan con micronutrientes de la dieta, como por ejemplo, el de la Metilentetrahidrofolatoreductasa (MTHFR). El gen de la MTFHR cataliza la reducción de la enzima THF a 5-metil THF (cosustrato para la remetilación de homocisteina a metionina). Si falla la enzima, aumenta la homocisteina en la sangre y hay mayor riesgo cardiovascular de trombosis. Este gen está instalado en el cromosoma 1, y consta de 11 exones. En el exon 4, en la posición 677, existe un polimorfismo que es debido a un cambio de citosina (C) por timina (T) (677 C/T). Ese cambio se traduce en un cambio de alanina por valina en el aminoácido 222 de la proteína final del gen, lo que conlleva menor actividad enzimática y mayor riesgo de cardiopatía coronaria. Se sabe, que en población general española, por ejemplo, las frecuencias de los distintos genotipos serían CC, CT, TT, 32, 52 y 16 %, respectivamente. Esa frecuencia tan elevada de T coincide con otros estudios realizados en poblaciones del sur de Europa y contrasta con la menor frecuencia del alelo T en el norte de Europa. Para esa variación se ha encontrado «una interacción gen-ambiente», con la ingesta de ácido fólico, vitamina del complejo B, presente en las verduras de hoja verde, frutas y legumbres. En sujetos con una dieta pobre en ácido fólico hay una mayor concentración sérica de homocisteina en los homocigotos TT, y un mayor riesgo cardiovascular. Sin embargo, en una dieta rica en ácido fólico se compensa el defecto en el ADN, y los individuos TT presentan una concentración sérica de homocisteina normal. Esto se observó, en el estudio de Framingham, por J.M. Ordovás (2007), que encontró mayor concentración de homocisteina asociada al genotipo TT en las personas con baja concentración de folatos (menor de 12.5 nanomoles por litro). Esa diferencia desaparecía cuando la concentración plasmática de folatos era superior.
Tal interacción gen-dieta explicaría la mayor prevalencia del genotipo TT en los países del Sur de Europa en comparación con los del Norte. En la Europa meridional, la dieta mediterránea, aporta cantidades elevadas de ácido fólico. Por ello, en el Mediterráneo las personas con el genotipo TT no tienen elevación sérica de homocisteina y evitan el riesgo cardiovascular. En el norte de Europa, la dieta aporta poco fólico, con elevación de homocisteina y del riesgo cardiovascular. Con el tiempo la selección natural, se habría encargado de reducir la frecuencia del genotipo TT en el Norte de Europa, pero eso ha determinado una mayor cantidad de muertes por cardiopatía coronaria.
5.2. La ventaja de los heterocigotos para una mutación nula en los Amish
Como ejemplo de «ventaja de los heterocigotos», encontramos lo detectado recientemente en las comunidades Amish, en el Condado de Lancaster (Pensilvania). Esta población es un grupo cultural y étnico fuertemente unido, descendiente de inmigrantes suizos de habla alemana. Desde que se formó en 1770, creen en el Nuevo Testamento de forma literal y se han aislado del mundo exterior, por lo que los 230.000 que se cree hay actualmente en Estados Unidos y Canadá, descienden de los mismos pocos cientos de fundadores del siglo XVIII. Es por tanto, una población con unas características ambientales (dieta y estilo de vida) especiales y rurales naturales, pues no han aceptado el desarrollismo de las sociedades occidentales. Al mismo tiempo poseen unas condiciones genéticas homogéneas en parte porque proceden de una pequeña población fundadora, y también porque practican desde hace muchos años cierta endogamia en sus uniones matrimoniales. Pero lo que es más interesante para lo que aquí nos ocupa, es que es un grupo de población donde las enfermedades cardiovasculares tienen una baja incidencia, lo que hasta ahora venía atribuyéndose a las condiciones ambientales y al tipo de vida natural que practican.
Sin embargo, recientemente se ha encontrado una «mutación nula», que no daría proteína, en el gen ApoC3 humano que controla la síntesis de triglicéridos precursores del LDLc -colesterol malo-, y de HDLc -colesterol bueno. En heterocigosis esta mutación parece conferir un perfil favorable de los lípidos plasmáticos, y por tanto un favorable efecto cardioprotector. Concretamente, se ha visto, que el 5% de los Amish de la región de Lancaster (EE UU), son portadores heterocigotos de esta mutación nula (R19X) en el gen que codifica la ApoC3 y como resultado expresan la mitad de la proteína con respecto a la presente en los no portadores. Los portadores presentan tanto en ayunas como en la etapa post-prandial -tras la ingesta- niveles muy altos de lipoproteínas de alta densidad (HDLc-colesterol bueno) y muy bajos de LDLc -colesterol malo-, con respecto a los no portadores. Esto significa un perfil global protector de CC, como se ha demostrado con los estudios de aterosclerosis sub-clínica medida por la calcificación coronaria. Todo ello sugiere un efecto protector de los heterocigotos para esta mutación, con respecto a los homocigotos mutantes y a los homocigotos normales no portadores de la mutación.
5.3. Pleitropía antagónica en proteínas de choque térmico y CC
Como ejemplo de pleitropía antagónica tendríamos el de la autoinmunidad. La inmunidad es el estado biológico caracterizado por tener suficientes defensas biológicas (humorales y celulares) para evitar la infección, la enfermedad, u otra invasión biológica no deseada. Entre las proteínas que actúan en la defensa inmunológica, están las chaperonas del tipo de proteínas del choque térmico (HPS), o proteínas de estrés.
Las HPSs tienen especial significación en la aterosclerosis por dos razones: 1º) porque representan la respuesta de las células de la pared arterial a varios estresantes, especialmente a los factores de riesgo clásicos, y 2º porque ellas son las dianas para las reacciones inmunes innatas y adaptativas, que inician y perpetúan el proceso inflamatorio vascular.
Así los primeros estadios de las lesiones ateroscleróticas, son clínicamente silentes hasta las etapas tardías de la vida. Cuando las células endoteliales arteriales aparecen estresadas por la persistencia de los diversos factores de riesgo clásicos, como ocurre en las edades post-reproductivas, expresan una inmunidad protectora a través de las HSP60 de las propias células alteradas, lo que promueve la aterosclerosis y desencadena el infarto de miocardio o cerebral. Por tanto, no se puede negar la importancia de los bien establecidos factores de riesgo, pero con esta teoría se les atribuye un nuevo papel en los estadios tempranos de la enfermedad, actuando como estresantes endoteliales.
Pero en el apartado que aquí nos ocupa -el de la pleiotropía antagónica- es de especial importancia en el contexto de la aterosclerosis una proteína de choque térmico concreta, la HSP60 que se localiza en el brazo largo del cromosoma 2 en la posición 33.1 (locus 2q33.1). Esta proteína es altamente inmunogénica aunque presenta gran conservación filogenética a nivel del ADN y de las proteínas. Por ejemplo, el promedio de identidad entre diferentes HPS60 bacterianas es del 90%, y de un 55% con la humana. Este hecho puede acarrear inesperadas reacciones cruzadas frente a infecciones bacterianas originando respuestas inmunes, humorales y celulares en la respuesta de las células autólogas humanas estresadas. Es decir, lo que se gana por un lado -en la defensa frente a los factores estresantes-, por otro se pierde -en las reacciones cruzadas frente a infecciones bacterianas.
Conclusiones
- Se cree que el acervo genético humano actual se moduló en gran parte en el Paleolítico. Este acervo genético, genotipo ahorrador por lo que se refiere a la alimentación, no se cambió en el Neolítico pues representaba una ventaja adaptativa, ya que afectaba a factores implicados en la lucha por la vida y la supervivencia (hasta la procreación). Actualmente las condiciones socio-culturales y el estilo de vida de las sociedades desarrolladas lo hacen especialmente incongruente, por lo que puede ser motivo del aumento de enfermedades como las cardiopatías coronarias.
- En aquellas poblaciones o regiones en las que se observa una mayor conservación y adherencia a dietas ancestrales (dieta mediterránea), hay una menor incidencia de enfermedades cardiovasculares, debido en parte a la interacción positiva de genes, como el de la MTFHR y la ingesta total del ácido fólico, procedente de la dieta.
- La baja frecuencia de enfermedades cardiacas de algunas poblaciones humanas como la de los Amish de EE UU puede deberse en parte a la relativa alta frecuencia en ellos de una mutación nula en genes que controlan factores lipídicos hemáticos de protección para el desarrollo de CC. Los favorecidos serían los heterocigotos con respecto a los dos tipos de homocigotos -normales y mutantes.
- La expresión de las proteínas de choque térmico o HSP, inductoras de aterosclerosis, como respuesta de las células endoteliales estresadas por los clásicos factores de riesgo, junto a la inmunorreactividad cruzada desarrollada a diversos microorganismos, es el coste que nosotros pagamos por tener una inmunidad protectora a diversos factores biológicos.
Luego, es posible que la aterosclerosis y su correlato pandémico las enfermedades cardiovasculares puedan tener, en parte, una explicación a la luz de la biología evolucionista, siendo una hipótesis susceptible de verificación y aplicable a la etiología y prevención de estos procesos.
Agradecimientos
Agradezco a los Dres. Manuel Ruiz Rejón y Rafael Navajas Pérez por su ayuda en distintas etapas de la redacción de este artículo.
Bibliografía general
- Campillo Álvarez, J.E. (2006). El Mono Obeso. Editorial Crítica.
- Canela, A., Vera, E., Klatt, P., y Blasco. M.A. (2007). High-throughput telomere length quantification by FISH and its application to human population studies. PNAS, 104(13):5300 – 5305.
- Fuster, V. (2008). La Ciencia de la Salud. Ediciones Planeta.
- Keys, M.A. (2007). Comer bien, sentirse bien y la dieta mediterránea. Editorial Ministerio de Agricultura.
- Nesse, R. y Williams, G. (2000). ¿Por qué enfermamos? Ediciones Grijalbo-Mondadori.
- Renaud, S. et al. (1992). Wine, alcohol, platelets and the French paradox for coronary heart disease. Lancet 339;1523-1526.